手賀沼流域における水生植物の現状
手賀沼流域における水生植物の現状
東京情報大学 齋藤康宏,富田瑞樹,原慶太郎
千葉県立中央博物館 林紀男
はじめに
湖沼や河川沿岸の植生帯は,水域と陸域の生態系をつなぐエコトーンとして重要な役割を果たしている.
たとえば,多くの魚類や水生昆虫,鳥類などに生息地や餌資源を提供することによって,高い生物多様性を維持している.
一般的には,生物多様性が維持されることでその生態系が健全に機能すると考えられており,その結果,水質浄化・
漁場提供・洪水制御などの多様な生態系サービスを提供するとされている.
近年,これらのサービスを資源として定量的に把握し,将来にわたって持続的に利用することが求められている.
千葉県北西部に位置する手賀沼は,我孫子市・柏市・印西市・白井市にまたがる面積6.5km2,
水量650万m3の浅い小湖沼である(図1).
手賀沼は水生植物が旺盛に繁茂する水域であったが,1960年代に戦後の食糧不足を補うために行われた国営干拓事業により,
植物が最も繁茂していた沼半分弱にもおよぶ529haの浅瀬が陸化し,多くの水生植物が絶滅・減少した.
その後の河川改修や,住宅団地や工業団地の造成にともなう生活排水・工業排水の増加,農業排水に含まれる
除草剤などが影響し,約30種近く生育していた在来水生植物のほとんどが消失してしまった.
一方,近年の手賀沼では,オオカナダモやオオフサモなどの外来水生植物が分布を広げつつある(林ほか 2008).
特に,環境省が特定外来生物に指定するナガエツルノゲイトウは流域内で急速に分布を拡大している(林ほか 2008)
ことから,在来種の保全や外来種の駆除の計画策定のために流域内における在来種・外来種の分布を把握し,
地理空間上にデータベース化することが重要である.
本報告では,千葉県立中央博物館および手賀沼水環境保全協議会の共催事業として市民と研究者との協働下で実施された
「手賀沼水草探検隊」によって得られた水生植物の分布図(林ほか 2008)を,GISを用いて地理空間上にデータベース化し,
手賀沼流域を構成する6つの流域における水生植物群集の種組成や空間分布を解析した結果から,
現在の手賀沼流域における水生植物の現状について報告する.
方法
手賀沼流域(156.2km2)は6つの流域から構成される(図2).小規模な谷津が入り組む亀成川流域には,
二次的自然が残存している.直接流入域は,手賀沼流域を構成しているすべての河川が流入してくる流域であり,
手賀沼・下手賀沼を含むため他の流域に比べ流域面積が広い.大津川流域には面積の大きい谷津に川幅の広い河川が流れ,
その下流域には水田がひろがる.大堀川流域の河川は市街地を流れ,その一部は暗渠となるなど,
流域の大部分が都市的環境である.金山落流域には区画整備された用水路が流れ,上流から下流まで水田がひろがる.
金山落流域と同様に,染井入落流域には区画整備された河川が流れるが,6流域の中で最も流域面積が狭い流域である.
水生植物の分布については,林ほか(2008)の手賀沼水草探検隊によって得られた水生植物の繁茂地点情報を,
GISを用いて地理空間上にデータベース化して解析に用いた.地形データ(10m DEM)から
河川の次数に対応する流域(以後,集水域とする)を作成し(図2),集水域の面積や河川の総延長を算出した.
集水域ごとに水生植物の在・不在を記録し,千葉県レッドデータブック掲載種か否か,および,
外来生物法による指定外来種か否かを調べた.ASTERデータ(2005年9月)から求めた正規化植生指数(NDVI)を用いて,
NDVI値が0.4以上のセルを植生域,それ以外を非植生域に区分したうえで,各流域における植生域の面積割合をもとめ,
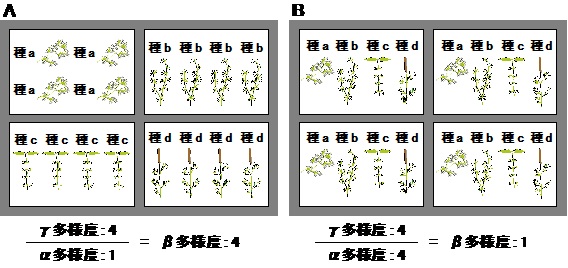
それを植被率とした(図3).種の多様性(豊富さ)を表す指標として,γ多様性,α多様性,およびβ多様性を用いた.
ある地域が複数の生育地からなる場合,全生育地の多様性をγ多様性,1つの生育地の多様性をα多様性,
生育地間の種組成の違いをβ多様性という(図4).本研究では,集水域ごとにGleasonの多様度指数を算出し,
平均したものを各流域のα多様度とした.各流域における最大次数の集水域の多様度指数をγ多様度とし,
β多様度をγ/αから算出した.
結果および考察
手賀沼流域に出現した21種のうち,約3割の6種が千葉県レッドデータブックに記載されている
保護指定種であったことから,貴重な水生植物が数多く生育することが確認された.
一方,外来生物法の指定種も6種と数多く出現していた.種数と多様度を流域ごとにそれぞれ比較すると,
亀成川流域には18種,直接流入域には17種出現し,γ多様度は5.65,4.57と,高い値を示した.
その他の流域におけるγ多様度は2未満と低かったことから,手賀沼流域における種多様性は,
亀成川流域と直接流入域によって引き上げられていると考えられた.すなわち,亀成川流域の
ような出現種の豊富な一部の流域が,手賀沼流域の水生植物の種多様性を高めていると推察された.
亀成川流域は流域面積が狭く,河川の総延長が短いものの,植被率は23.3%と他流域よりも顕著に高かった.
また,α多様度が3.10と他流域に比べて非常に高い値を示した.一方,大津川流域と大堀川流域は,
それぞれの流域面積を合計すると手賀沼流域の約4割を占めるものの出現種数は6種や4種と少なく,
α多様度はどちらの流域も1未満(0.83と0.40)と低い値を示した.また,植被率もそれぞれ3.7%,1.9%と低かった.
手賀沼流域の西部は1960年代から土地開発が進んでいるために市街地面積割合が高い一方,
流域東部は市街地化の影響が低いため二次林などに由来する緑地面積の割合が高かった(図3).
すなわち,東西の開発程度の違いが水生植物の種多様性の差異に強く影響しているものと推察された.
亀成川流域には,ツツイトモやイトモなどの千葉県レッドデータブック指定の沈水性保護生物が多数確認された.
沈水性の水生植物は水底に固着しながら植物体全体が水中にあるため,
他の生活型よりも水質や光環境の影響を受けやすい.つまり,宅地開発や森林伐採による泥水の流入,
宅地の増加に伴う生活排水の増加,ホテイアオイなどの外来性浮遊植物の繁茂によって水中へ届く光が大きく減少することで,
沈水植物は急速に消滅してしまうことが予想される.また,保護生物の分布は,東西による開発程度の違いに加え,
亀成川流域の谷津の多さにも影響されていると考えられた.
すなわち,より小さい谷津が複雑に入り組んだ樹枝状の地形を示す亀成川上流部に残存する自然環境が,
保護生物に区分される水生植物の生育を維持しているものと推察された.
また,一部の水生植物は生育場所によって生活形態が変わる.たとえば,水深の確保された場所では,
沈水植物として知られるササバモは,水中葉が乾出すると沈水葉が浮葉に形態変化する.
さらに水位が低下し植物体が乾出する条件になると新たに陸生葉を形成し湿生植物になる.
これらのような,水面に葉を広げる浮葉植物や,水面を浮遊する浮遊植物は流速の速い水域には定着できないため,
ため池などの閉鎖性水系やワンドなど止水域の存在が重要である.直接流入域には手賀沼・下手賀沼が,
亀成川流域にはため池などがあることが,これらの流域において水生植物の多様度が高い要因のひとつと考えられた.
一方,市街地が流域面積の多くを占める大津川流域・大堀川流域・染井入落流域では,
ヨシやマコモといった抽水植物が主に確認された.抽水植物は,沈水植物と同じく水底に根をはるが,
植物体の一部(茎や葉など)を空気中に出す.このため抽水植物は,好適な光環境に起因して生産性が高く,
人間活動による汚濁の進んだ水系でもよくみられることが知られている.本研究結果でも同様の傾向が確認されたといえよう.
手賀沼流域における水生植物群集の種組成や空間分布は,多様な生育環境(河川・ため池・湿地・水田)の有無や,
過去の宅地造成,河川改修によるハビタットの喪失などの直接的影響,その後の水質の変化,
すなわち,農業排水・生活排水などによる間接的影響に大きく影響されながら形成されてきたものと考えられた.
現在,土地開発の影響が比較的少なかった亀成川流域においても2009年から河川改修工事が本格化していること,
特定外来生物のナガエツルノゲイトウが手賀沼流域南東の印旛沼水系から農業用水路を介して侵入し,
手賀沼流域全体に分布を広げた(林ほか 2009)ことなどにより,
手賀沼流域における在来水生植物群集は依然として危機的状況にある.本研究結果が,
今後の保全・復元活動の一助になれば幸いである.
引用文献
林紀男・谷城勝弘・浅間茂 (2008) 手賀沼流域における水生植物の繁茂状況. 千葉生物誌 58(1):1-6
林紀男・横林庸介・竹中真里子 (2009) 手賀沼流域におけるナガエツルノゲイトウの繁茂域の変遷. 水草研究会誌 91:6-10
この小文は日本景観生態学会第19回新潟大会で発表したものをとりまとめたものである.
図1
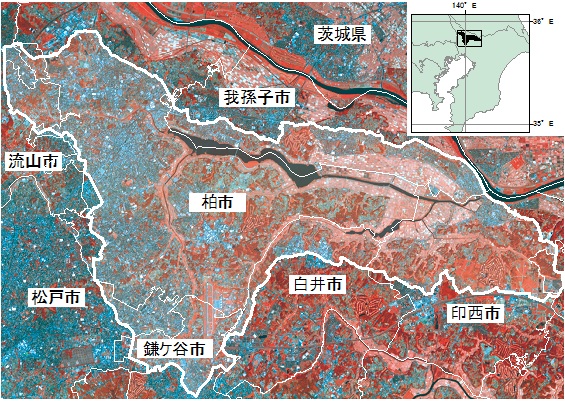
図2
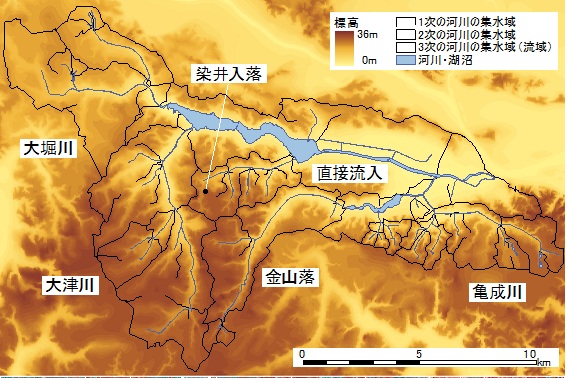
図3
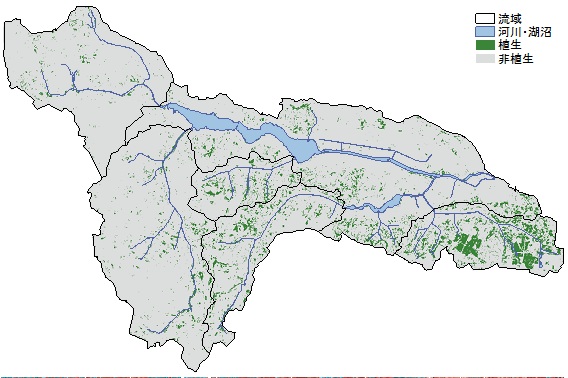
図4