 日本列島と中国との対比―植生の視点から
日本列島と中国との対比―植生の視点から
千葉県立中央博物館 原正利
ユーラシア大陸東縁部は,亜熱帯においても乾燥気候が卓越することなく,熱帯から寒帯まで湿潤な気候が連続する世界で唯一の地域である.このことは,植生的には,森林植生が緯度に沿って連続的に推移し,途中を草原や沙漠によって分断されないことを意味している.この地域では,常緑性の熱帯林と夏緑性(落葉性)の温帯林の間に,常緑性であるが様々な点で熱帯林とは明らかに区別される森林,すなわち照葉樹林あるいは亜熱帯・暖温帯常緑広葉樹林と呼ばれる森林が広く分布している.この森林の優占種はクスノキ科やブナ科の樹木で,クスノキ科-ブナ科林Lauro-Fagceous forestとも呼ばれている.
植物や植生の分布を支配する最も重要な気候要因は,気温と降水量であるが,ユーラシア大陸東縁付近では,後者は十分にあるため,気温のみが支配要因となる.温度は2つの側面から植物の生育に影響する.ひとつは,光合成による物質生産量を支配する面である.光合成は酵素を介した化学反応である.したがって,酵素が失活する一定の高温までは,光合成速度は,一般に温度に比例して増加する.さらに,季節によって気温が変化する状況を想定すると,1年間の総光合成量は季節ごとの光合成量の積分値として与えられることとなる.これを温度の面から指標化したものが有効積算温度で,様々なものがあるが,日本列島では吉良(1945)の考案した暖かさの指数WIが広く使われている.
温度要因が植物の生育に影響するもうひとつの面は,低温すなわち寒さが,凍結などを介して植物体に直接的ダメージを与えたり,受精や花粉管の伸張など生活環の一部を阻害する場合である.庭に植えた園芸植物が霜に当たって一晩のうちに枯れてしまうことは,多くの人々が経験するところである.また,常緑広葉樹の分布の北限は,冬の低温によって決定され,最寒月平均気温-1℃と一致すると言われている.吉良(1949)は寒さの指数CIという指標も考案していて,これによればCI=-10℃から-15℃が常緑広葉樹林帯の北限と一致する.日本列島では,WI=45~85℃・月(ただしCIが-10℃以上の範囲)が冷温帯夏緑広葉樹林,WI=85~180℃・月が暖温帯常緑広葉樹林,WI=180~240℃・月が奄美大島以南の亜熱帯常緑広葉樹林に対応とされている.
本研究プロジェクトでは,植生帯ごとに,植生の分布や動態を明らかにしていくことを目指している.そのためには,日本列島の植生帯と中国本土の植生帯を対比して対応関係を明確にしておく必要がある.上記のような気候条件を解析すれば,このことは,一見,容易に果たせるように思われるが,実際には様々な点を考慮しなければならず一筋縄ではいかない.
まず,大陸と島という地形の違いを反映した環境条件の違いがある.図1に東アジアにおけるWIおよびCIの等値線の分布を示す.この図から,東アジアの中でも,日本列島~台湾に至る島嶼部と中国本土では,等値線の走行に緯度的なずれがあり,島嶼部では等値線が北上していることがわかる.例えばWI=180℃・月のラインは大陸部では福建省の沿岸部をかすめるように通っているが,日本列島では北緯約30°,屋久島と奄美大島の間を通っている.黒潮の影響もあって,同緯度で比較した場合,日本列島のほうが温暖なのである.一方,WI=140℃・月のラインは大陸部では浙江省の北部をとおり,日本列島でもそれほど北上せずに九州南端部をかすめている.結果的に,中国本土で緯度的に広い範囲を占めるWI=140~180℃・月の範囲は,日本では屋久島など,ごく狭い地域にしか存在しないこととなる(Kira, 1991).
当研究プロジェクトで重要な調査地のひとつとしている天童山は,ちょうど,WI=140℃・月のライン上に位置している(図1).ここには発達した照葉樹林が残されている(図2).森林の構成種を見ると,宮崎県など九州南部の照葉樹林と比べても,はるかに種が豊富である.例えばブナ科は19種,クスノキ科は18種が記録されている(宋・王,1995).また,日本では奄美大島以南(WI > 180℃・月)に限って分布するため,亜熱帯の指標種のように考えられているイジュSchima superba(日本のイジュSchima wallichiiと同種または極めて近縁な種,ツバキ科)も分布している.すなわち,日本と比べると,WIの値とフロラやその多様性にずれがある.これはなぜだろうか.
日本列島では,常緑広葉樹林は,四国や九州などのWI=85~140℃・月の範囲に,最も広く分布している.一方,中国ではこの範囲には,常緑広葉樹林は分布するものの連続的でなく,夏緑広葉樹のほうが優勢である.常緑広葉樹林は,主に揚子江以南に限られる.気温的には,日本よりもかなり高く,WI=130~140℃・月以上の範囲である.これは,単に低温だけでなく,乾燥によって,常緑広葉樹林の北限が南に押し下げられているためと考えられている(Fang et al., 1989, 2002).天童山は中国では常緑広葉樹林の北限近くに位置するにもかかわらず,日本と比べて種の多様性が高いのは,常緑広葉樹林の北限を決定している要因に違いがあり,常緑広葉樹フロラとしては,屋久島以南に相当する豊かな内容を,そのまま保持しているためと考えられる.
さらに,日本列島では地史も森林の種多様性に大きく影響している.もともと日本列島は大陸の一部であった陸塊が,日本海の拡大によって1600万年前頃までに大陸から切り離されて形成されたが,その後も中国本土と連続していた時期も多く,日本列島の生物相は大陸からの生物の移動と絶滅を通して形成されてきたものである.しかし,島となってからは,海峡によって移動を妨げられることも多く,生物相が大陸と比べて不調和になっている面がある.例えば,奄美大島以南の南西諸島は,WI=180℃・月以上の亜熱帯にあり,日本ではこの地域にしか分布しない亜熱帯性の植物群を豊富に有しているが,一方,森林の上層を形作っている樹木はスダジイCastanopsis sieboldiiの亜種であるオキナワジイCastanopsis sieboldii subsp. lutchuensisやアラカシQuercus glaucaの変種であるアマミアラカシQuercus glauca var. amamiana,タブノキMachilus thunbergiiなど九州以北の常緑広葉樹林と共通する種が大半である(図3).これは島であるために,ブナ科のように種子散布力の小さな植物は侵入することができず,本来の亜熱帯性植物相の形成が妨げられたためと考えられている(堀田,1974;吉良,1989).逆に,イジュSchima wallichiiが奄美大島以南に限られ,九州南部や屋久島に見られないのも地史的な理由が関係しているかもしれない.
中国と日本の植生タイプの違いは,夏緑広葉樹林についても見られる.日本列島では冷温帯の森林植生はブナ林によって代表される(図4).一方,中国では,冷温帯の森林植生はブナ林ではなく,モンゴリナラQuercus mongolicaなどのナラ類や,カエデ科,シナノキ科,カバノキ科などの落葉広葉樹が主体の森林である.それも主に東部に限られ,西部では見られない.中国にもブナ属が優占する森林はあるが,常緑広葉樹林帯の山岳に点在して分布するだけで成帯性の植生とはなっていない.日本列島の冷温帯域でブナ林が卓越した分布を示すのは,大陸と比べて湿潤であること,特に,積雪量が多いため,冬から春先の時期も乾燥しないことが関連していると考えられている.
以上のように,中国と日本は,共に東アジアに位置しながら,大陸と島という地理的条件の違いがあり,これに起因して,気温や降水量など気候要因の組み合わせが違い,また地史も異なる.このため,植生やその動態を考える上で,単純な比較は難しい.両国の植生を正確に対比した研究は,まだ,無いのが現状である.本研究プロジェクトでは,衛星画像に基づく鳥瞰的な視野を持ちつつも,蟻の眼を持って植生の内容を正確に把握し,対比していくことが必要であると考えている.
----図の説明-----
図1.東アジアにおける暖かさの指数WIおよび寒さの指数CIの等値線の分布.Kira(1991)に基づく.
図2.中国浙江省天童山の森林.名刹天童寺を取り囲むように,照葉樹林(常緑広葉樹林)の自然林が残されている.九州南部の照葉樹林と比較しても種の多様性は高い.
図3.沖縄島ヤンバルの森.ヤンバルクイナも暮らす森には,アカネ科やヤブコウジ科など亜熱帯の植物が多く見られるが,森林の上層は,主にブナ科のオキナワジイCastanopsis sieboldii subsp. lutchuensisとオキナワウラジロガシQuercus miyagiiによって構成され,単純である.
図4.奥羽山地のブナ林.日本列島の多雪山地にはブナ林が広く分布する.宮城県栗駒山.
図1
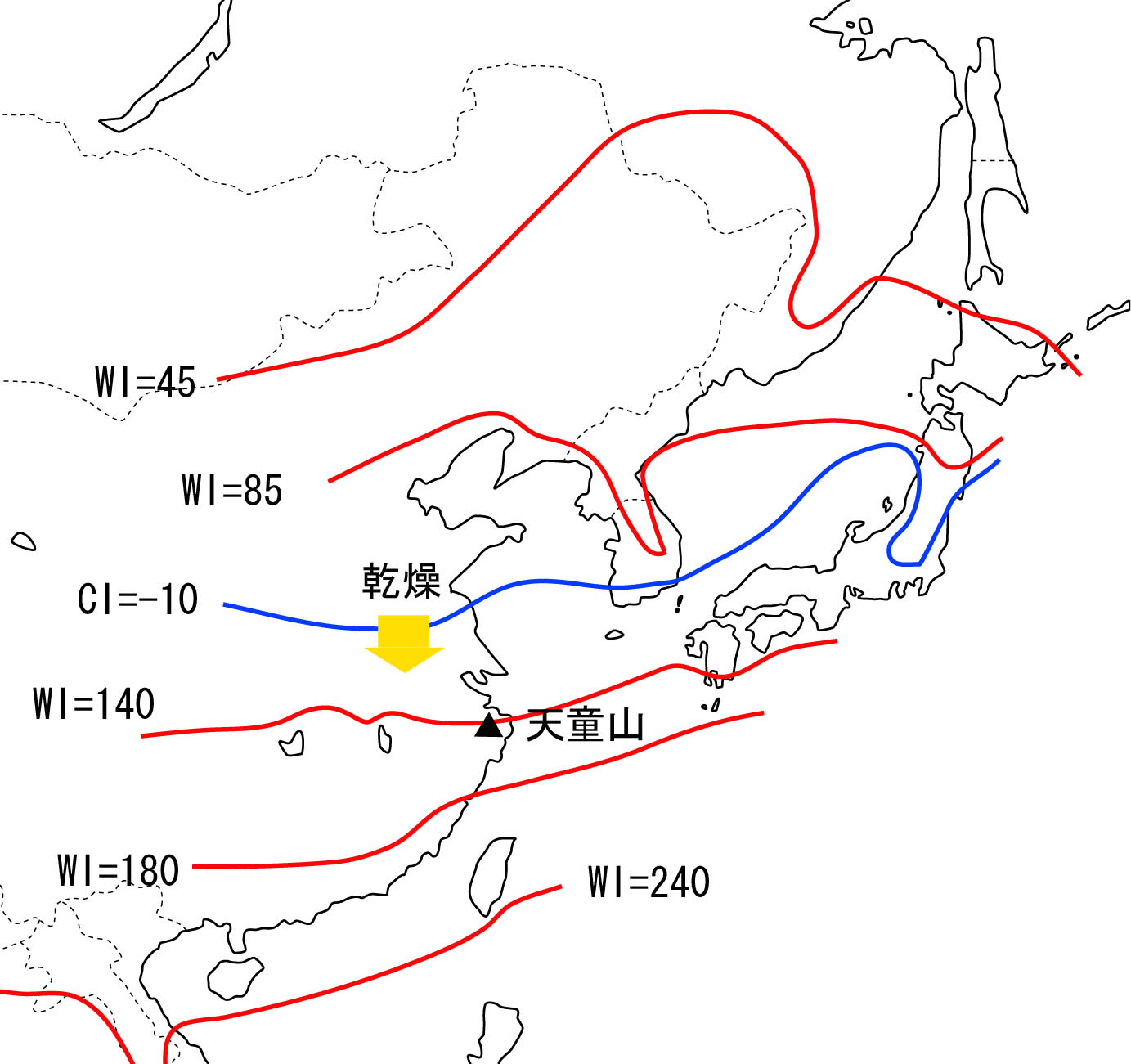
図2

図3

図4


